


 雷达卡
雷达卡



本文内容速览:
自交不亲和性的基本概念
与人类避免近亲结婚以降低隐性有害基因纯合风险类似,植物作为固着生物也面临因自花授粉引发的近交衰退问题。为了防止此类情况,许多被子植物在进化过程中发展出一种称为自交不亲和性(Self-incompatibility, SI)的生殖机制。研究表明,至少有100多个被子植物科存在这一现象(Boris et al., 2008)。SI是指植物虽能形成正常的功能性雌雄配子,但在进行自花授粉或授粉后无法完成受精结籽的过程。
对植物而言,是否具备自交不亲和性是一项关键的进化策略选择。若为自交亲和,植物可在低交配成本下迅速固定优良基因型,利于种群快速扩张和定殖;但其遗传基础狭窄,抗环境胁迫能力弱。相反,自交不亲和虽需依赖外部传粉媒介、提高繁殖成本,却有助于维持较高的遗传多样性,增强对多变环境的适应力,并促进种内及种间生殖隔离,从而在长期演化中具有显著优势。
自交不亲和植物的生殖特征差异
相较于自交亲和植物,自交不亲和类型在空间结构、发育时序以及传粉方式上表现出明显区别。从空间角度看,这类植物常将雄蕊与雌蕊分布于不同个体或花序中,以物理方式规避自交。在时间维度上,则通过雌雄器官异时成熟——即雄蕊与雌蕊不在同一时期达到可育状态——进一步减少自花授粉可能。而在传粉机制方面,多数依赖昆虫或其他动物媒介传播花粉,借助多样化的访花者降低同株授粉频率。
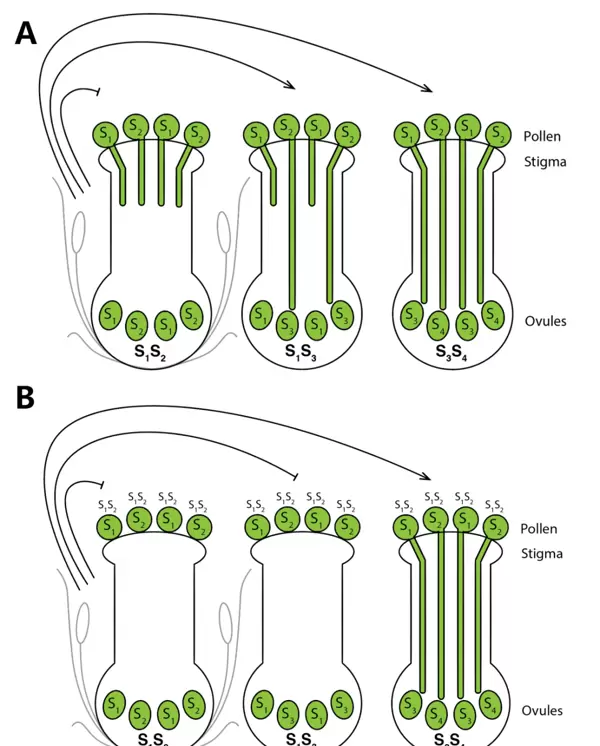
GSI与SSI的作用模式解析
在同型自交不亲和系统中,主要分为配子体自交不亲和(Gametophytic self-incompatibility, GSI)和孢子体自交不亲和(Sporophytic self-incompatibility, SSI),二者在识别机制与反应位点上存在显著差异。
在GSI系统中,花粉的亲和性由其自身单倍体基因型决定,不受二倍体亲本影响。当花粉携带的S单体型与胚珠相同时,表现为不亲和。此时花粉通常能够正常水合、萌发并穿透柱头,但在花柱内部花粉管会破裂,导致无法抵达子房完成受精。
而在SSI系统中,花粉是否被接受取决于产生它的二倍体亲本(孢子体)的S基因型。该过程主要发生在柱头表层:一旦花粉带有与柱头相同的S等位基因,柱头乳突细胞表面便会沉积胼胝质,阻止花粉萌发或穿透。
自交不亲和的分类及其遗传机制
根据花朵形态差异,开花植物中的自交不亲和可分为异型自交不亲和(Heteromorphic self-incompatibility, HSI)和同型自交不亲和(Homomorphic self-incompatibility)。HSI可通过肉眼观察到花部结构的不同,如同株具长短柱花;而同型种类在外形上无明显区别,只能通过遗传分析判断。
自交不亲和的核心在于植物对“自我”与“非自我”花粉的识别能力,这一过程通常由一个高度多态性的S位点控制(少数物种涉及两个或多个S位点)。该S位点包含紧密连锁的雌蕊决定因子和花粉决定因子,共同构成一个完整的遗传单元——S单体型。当雌蕊与花粉拥有相同的S单体型时,触发信号级联反应,抑制自体受精;反之,若S单体型不同,则允许正常受精进程。
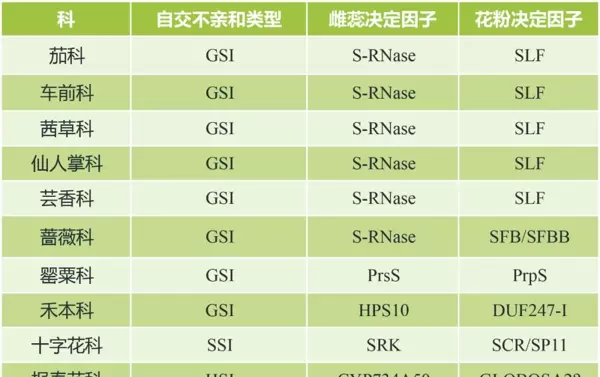
表1 不同植物的自交不亲和类型(信息来源:Zhang et al., 2024)
注:表中列出的是各科代表性物种的主要自交不亲和机制,实际分类中可能存在例外情况,具体应以物种为单位进行分析。“?”表示当前研究尚未完全确认相关因子。
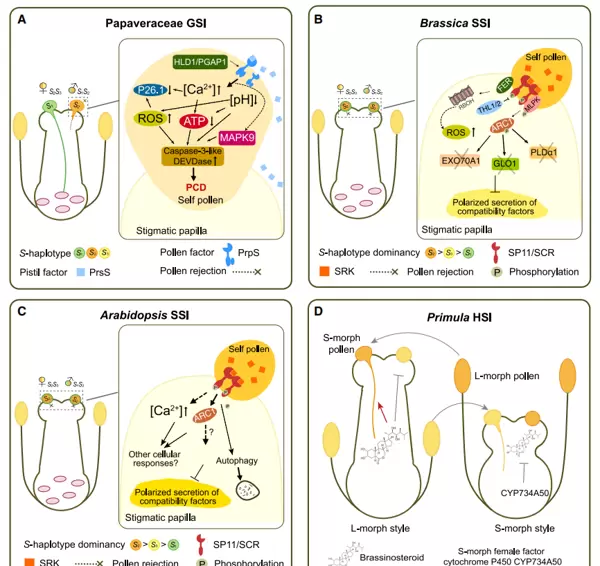
图2 展示了不同自交不亲和系统的分子机制(Zhang et al., 2024)。
(A)在罂粟科的配子体型自交不亲和(GSI)系统中,花粉决定因子PrpS与雌蕊分泌的PrsS蛋白发生特异性识别,触发Ca2+依赖性信号通路。该过程引起细胞质内游离Ca2+浓度升高、活性氧(ROS)积累、ATP合成减少、细胞质酸化,并抑制可溶性无机焦磷酸酶P26.1的活性。同时,MAPK9和类caspase-3/DEVDase等SI-细胞程序性死亡(SI-PCD)关键调控因子被激活,最终诱导自花花粉发生程序性死亡。
(B)在十字花科芸薹属的孢子体型自交不亲和(SSI)系统中,SRK受体激酶与MLPK共磷酸化后激活E3泛素连接酶ARC1,进而介导对花粉兼容相关蛋白EXO70A1、GLO1和PLDα1的泛素化降解。此外,FER-Rac/Rop-Rboh信号模块也被激活,促进NADPH氧化酶大量产生活性氧(ROS),从而迅速排斥不亲和花粉。
(C)拟南芥属同样属于十字花科SSI系统,其机制涉及SRK与小肽SCR的相互作用,引发柱头乳突细胞中胞质Ca2+快速上升并激活自噬途径,最终导致花粉兼容因子功能受抑,实现对自花花粉的排斥反应。
(D)报春花科的异型自交不亲和(HSI)表现为长花柱型(柱头高于花药)与短花柱型(柱头低于花药)两种表型。其中,短花柱型由细胞色素P450家族成员CYP734A50调控,该基因通过抑制油菜素内酯的生物合成,阻碍细胞伸长,从而使花柱缩短,参与SI表型的建立。
配子体型自交不亲和(GSI)是自然界中最广泛存在的SI类型,主要分为两类:一类是以S-RNase为基础的核酸酶系统,另一类则是以SI-PCD为核心的程序性死亡系统。
在茄科、车前科、茜草科、仙人掌科、芸香科及蔷薇科植物中,GSI由雌蕊端的S-RNase蛋白主导。当授粉亲和时,S-RNase在花粉管中被有效降解或失活,允许花粉管正常生长;而在不亲和情况下,S-RNase保持活性并降解花粉管中的RNA,阻碍其延伸。
相比之下,罂粟科采用的是SI-PCD型GSI机制。其雌蕊决定因子PrsS为一种分泌至柱头表面的蛋白质,而花粉决定因子PrpS则是一种跨膜受体。二者特异性结合后,在不亲和授粉条件下引发细胞内Ca2+波动和ROS爆发,启动多层级信号级联,最终导致花粉细胞程序性死亡(见图2A)。
禾本科植物的GSI机制更为特殊,由两个非连锁的多等位基因位点 S 和 Z 共同控制。其中, S 位点编码一个雌蕊因子sS以及两个花粉因子SDUF247-I和SDUF247-II; Z 位点则表达一个雌蕊因子sZ和两个花粉因子ZDUF247-I和ZDUF247-II。只有当花粉携带的 S 与 Z 等位基因均与雌蕊完全匹配时,才会触发自交不亲和反应。
除GSI外,十字花科中的芸薹属和拟南芥属表现出典型的孢子体型自交不亲和(SSI),其雌蕊决定因子为丝氨酸/苏氨酸受体激酶SRK,花粉决定因子为小肽SCR(又称SP11)。在不亲和授粉过程中,SRK与SCR结合后在柱头细胞中引发一系列磷酸化事件,导致下游兼容因子被抑制,从而阻止自花花粉萌发与侵入(如图2B、C所示)。
此外,还存在异型自交不亲和系统(HSI),典型代表为报春花科植物。这类系统依赖于花部结构的空间分离,其花柱长度多态性与SI特性紧密连锁。CYP734A50作为关键调控基因,通过降低油菜素内酯水平抑制细胞伸长,促成短花柱形态的形成,进而影响授粉结果(见图2D)。
自交不亲和机制在作物育种中具有重要应用价值。目前生产F1代杂交种的主要技术路径包括自交不亲和系和雄性不育系两种方式。利用自交不亲和系作为母本与其他品系(如其他不亲和系、自交系或常规品种)杂交,具备操作简便、亲本繁殖容易、杂种优势显著以及制种成本较低等优点。
例如,20世纪70年代我国科研人员基于十字花科植物的自交不亲和特性,成功培育出甘蓝型杂交油菜新品种,显著提升了产量和品质。此外,该机制也在甘蓝、大白菜、萝卜等多种蔬菜的杂交育种中得到广泛应用。
然而,在实际育种过程中也面临挑战:F1代杂交种所使用的亲本若自交不亲和程度较高,则难以进行自身繁殖。为此,研究者提出了多种克服策略,包括采用物理或化学手段处理柱头以暂时解除不亲和反应,或通过遗传工程干扰、敲除关键SI基因来获得稳定的自交亲和材料(参见图3)。
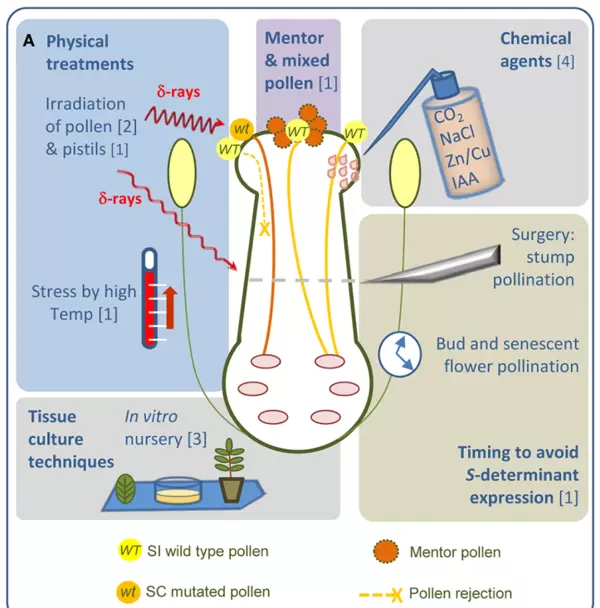
图3 概括了打破自交不亲和性的多种策略(Muoz-Sanz et al., 2020):
- 物理方法:对花粉或雌蕊组织施加辐射或高温处理;
- 花粉蒙导效应:将亲和花粉(蒙导花粉)与不亲和花粉混合授粉,借助其引导作用促进后者萌发;
- 化学试剂处理:使用特定化合物干预信号传导;
- 抑制S决定因子表达:通过基因沉默等方式阻断SI关键基因的功能;
- 组织培养技术:结合基因编辑手段,靶向干扰或敲除植株中的自交不亲和相关基因,创制自交亲和新种质。
在现代农业生物技术中,基因编辑技术为调控自交不亲和性提供了精准工具。主要有两种应用场景:
第一,在原本自交不亲和的物种中,通过编辑SI关键基因(如SRK、SCR、S-RNase等),破坏其功能,从而构建出自交亲和的稳定品系,便于种子扩繁和杂交育种操作。
第二,在通常自交亲和的物种中,可通过编辑那些抑制SI通路的负调控因子,重新“唤醒”潜在的自交不亲和机制,构建人工SSI系统,用于维持杂种优势或防止基因漂移。
这两种策略不仅拓展了作物育种的技术路径,也为未来设计更高效、可控的杂交制种体系提供了理论基础与实践方案。
马铃薯的育种与生产长期以来主要围绕四倍体类型展开。由于其基因组高度杂合、遗传背景复杂,导致育种周期较长,品种更新速度缓慢。此外,马铃薯依赖块茎繁殖,带来了较高的储运和脱毒成本。为突破这些瓶颈,欧洲多家育种机构早在十余年前便启动了二倍体马铃薯的育种研究。在我国,由中国农业科学院黄三文研究员牵头,联合多方单位于2017年正式启动“优薯计划”,旨在构建高效的二倍体马铃薯杂交育种体系。
要实现这一目标,首先需建立稳定的高代自交系群体,以确保杂交后代的一致性。然而,大多数二倍体马铃薯具有自交不亲和性,严重制约了纯合系的培育。根据表1信息可知,马铃薯属于配子体型自交不亲和(GSI)系统,其中 S-RNase 基因是决定雌蕊特异性的关键因子。
2018年8月,中国农科院黄三文与张春芝团队在《Nature Plants》上发表了题为“Generation of self-compatible diploid potato by knockout of S-RNase”的研究成果。该研究利用CRISPR/Cas9基因编辑技术靶向敲除 S-RNase 基因,使其编码的蛋白失去功能,成功获得了自交亲和的二倍体马铃薯材料。这项突破为后续高效育种提供了重要基础,显著提升了马铃薯遗传改良的潜力。
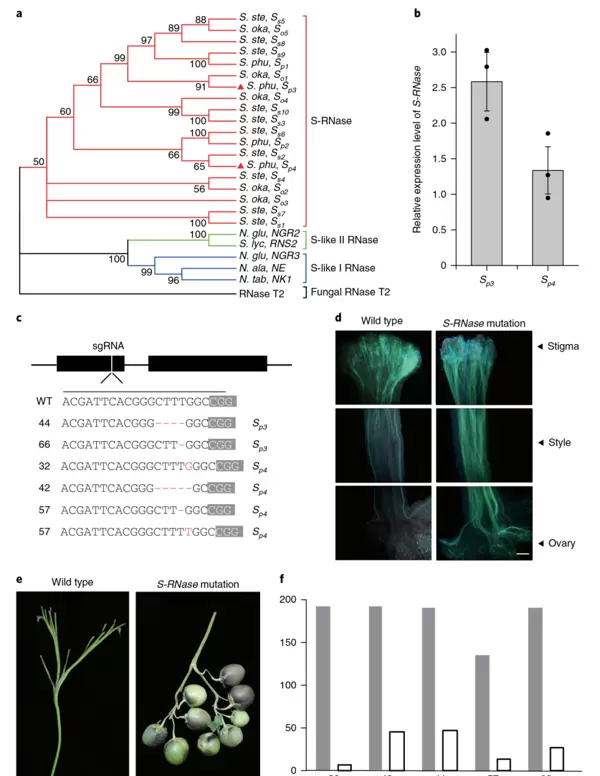
图4 敲除 S-RNase 基因克服马铃薯自交不亲和性(Ye et al., 2018)。 (a)不同 S-RNase 基因的进化关系分析; (b)在二倍体马铃薯S15-65材料花柱中两个 S-RNase 等位基因的表达水平; (c)S-RNase 基因结构及编辑植株中的突变类型; (d, e)通过苯胺蓝染色观察野生型与敲除植株自交授粉后花粉管生长情况及坐果表现; (f)T0代自交产生的T1代植株中 Cas9 基因分离情况检测,灰色表示检测总数,白色代表不含 Cas9 的植株(Cas9-free)。结果显示:野生型植株花柱中花粉管生长受阻,停滞于花柱上半部,无法结实;而敲除植株中花粉管可顺利延伸至胚珠,并形成饱满果实。同时,T1代检测证实 S-RNase 突变可稳定遗传,并能获得无外源基因残留的Cas9-free个体。
在其他作物中,调控自交不亲和性的研究也取得重要进展。例如甘蓝型油菜虽为自交亲和物种,但其祖先白菜和甘蓝均为自交不亲和类型。农业生产中常借助自交不亲和机制进行杂交育种。若希望重新创制自交不亲和的甘蓝型油菜,传统方法包括种间杂交或将白菜中的 BrSCR 基因导入,但前者后代不稳定且需多轮回交,后者涉及外源基因插入,难以获得非转基因品种,因此目前可用的自交不亲和油菜品种极为有限。相比之下,CRISPR/Cas9基因编辑技术提供了一条精准高效的替代路径。
2021年5月,华中农业大学马朝芝与戴成课题组在《Plant Biotechnology Journal》发表题为“Generation of novel self-incompatible Brassica napus by CRISPR/Cas9”的论文。研究推测,BnS6-Smi2 基因可能抑制了 BnSCR7 的表达,从而导致自交亲和。通过CRISPR/Cas9技术敲除 BnS6-Smi2,研究人员成功恢复了 BnSCR7 的活性,进而创制出新型自交不亲和的甘蓝型油菜材料。
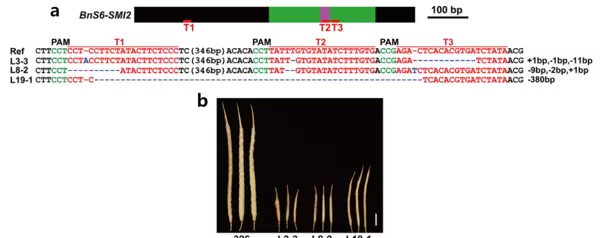
图5 利用敲除 BnS6-Smi2 基因创制自交不亲和甘蓝型油菜(Dou et al., 2021)。 (a)BnS6-Smi2 基因结构、序列特征及突变形式; (b)野生型326与T1代敲除株系SI-326自花授粉8周后果荚表型对比。结果表明,三个SI-326株系的果荚明显缩短,部分为空荚,进一步验证该现象由自交不亲和引起。
近年来,随着对自交不亲和分子调控机制的深入解析,科研人员不仅能够定向创制新种质,还拓展了对远缘杂交中种间、属间生殖隔离机理的理解。多项高水平研究成果相继发表于国际顶级期刊。
2021年4月,华东师范大学李超课题组在《Science》发表题为“PCP-B peptides unlock a stigma peptide-receptor kinase gating mechanism for pollination”的研究。该工作以拟南芥为模型,揭示了花粉表面的PCP-Bγ小肽如何竞争性干扰柱头中的RALF33小肽,进而抑制RALF33-FER/ANJ受体激酶通路所维持的活性氧(ROS)水平,最终影响花粉水合与萌发过程,阐明了柱头-花粉识别的关键分子机制。
2023年1月,山东农业大学段巧红团队与美国马萨诸塞大学Alice Y. Cheung课题组合作,在《Nature》发表相关研究,进一步深化了对该类信号传导网络的认知,推动了植物生殖生物学的发展。
2023年10月,北京大学瞿礼嘉与钟声课题组在《Cell》杂志发表题为“Antagonistic RALF peptides control an intergeneric hybridization barrier on Brassicaceae stigmas”的研究论文。该研究从分子层面揭示了拟南芥柱头如何识别并接纳同种花粉,同时排斥远缘花粉的机制。研究人员提出了柱头与花粉之间相互识别和信号传递的“锁-钥模型”,系统阐明了十字花科植物在柱头部位形成种间及属间生殖隔离的机理。该模型不仅合理解释了长期存在的“花粉蒙导效应”,还通过外源施加人工合成的pRALF小肽——相当于“钥匙”——成功打开柱头上的“锁”,使远缘花粉得以穿透柱头,突破生殖屏障,最终实现了不同属间的远缘杂交胚获得。
同期,另一项重要研究发表于《Nature》杂志,题为“Stigma receptors control intraspecies and interspecies barriers in Brassicaceae”。该研究发现十字花科植物通过调控柱头活性氧(ROS)水平来维持种内和种间的生殖隔离。研究团队通过外源应用ROS清除剂(如Na-SA)、一氧化氮供体(GSNO),或采用反义寡核苷酸(AS-ODN)技术干预相关通路,成功获得了大白菜的种间乃至属间远缘杂交胚胎。这一成果为植物远缘杂交育种及新种质资源创制提供了全新的策略与理论支持。
对于主要研究自交亲和物种(如水稻、大豆、玉米等)的研究者而言,“自交不亲和”现象可能较为陌生,甚至难以理解其生物学意义。然而,自交不亲和性在自然界中广泛存在于多种植物中,尤其在十字花科、茄科和蔷薇科中尤为普遍。近年来,围绕该现象的分子机制研究持续升温,取得了多项突破性进展。本文简要梳理了自交不亲和的基本概念与拓展知识,并重点介绍了上述两项关键研究成果,旨在帮助读者建立必要的背景认知,从而更深入地理解相关领域前沿论文的核心内容。

 京公网安备 11010802022788号
京公网安备 11010802022788号







