


 雷达卡
雷达卡



冰冻圈涵盖了地球上所有以固态水形式存在的区域,包括极地和高山地区的冰川、冰盖、多年冻土以及海冰等。尽管低温环境对生命构成严峻挑战,这些寒冷生态系统却蕴藏着丰富且独特的微生物多样性。这些微生物不仅是极端环境下生命适应能力的典范,更是驱动冰冻圈内关键生物地球化学过程的核心力量。它们在碳、氮、硫等元素循环中发挥着不可替代的作用,支撑着整个生态系统的运行。
近年来,随着宏基因组学、宏转录组学等高通量分子技术的广泛应用,科学家得以深入揭示冰冻圈微生物群落的组成结构与功能潜力。研究发现,这些微生物演化出了多种冷适应机制,如合成抗冻蛋白、调整细胞膜脂质组成、优化低温酶活性等,使其能够在接近或低于冰点的环境中维持代谢活动。无论是被封存在古老冰层中的休眠细胞,还是活跃于融雪期土壤表层的功能类群,都展现出惊人的生存策略与生态韧性。
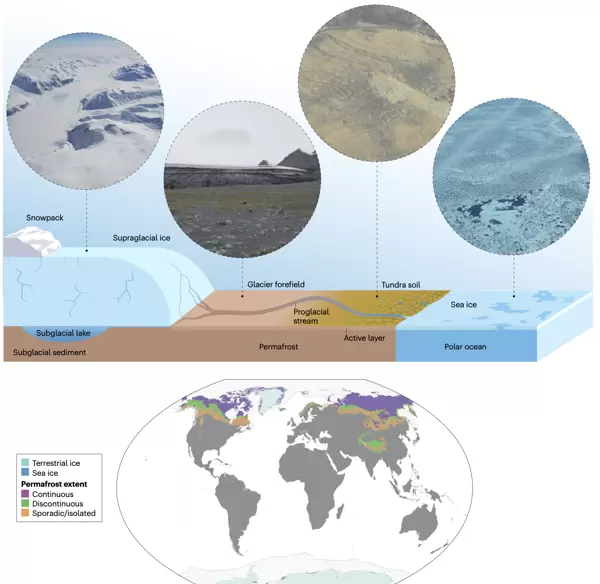
一个概括性极地生态系统的示意图和照片,其中包括冰盖、冰川及相关的冰川前缘、附近的多年冻土与活性层土壤,以及有海冰覆盖的海洋。这些生态系统共同覆盖了地球表面的相当大部分。地图显示了全球冰盖和冰川的分布、海冰中位最大范围(1981-2010年)以及连续(覆盖90-100%的景观)、不连续(覆盖50-90%的景观)和零星(覆盖<50%的景观)多年冻土的分布范围。
然而,气候变化正以前所未有的速度重塑冰冻圈的物理格局。特别是在北极地区,变暖幅度达到全球平均水平的两倍以上,这种“极地放大效应”引发了广泛的环境剧变:冰川和冰盖持续退缩,海冰覆盖时间缩短、厚度减薄,多年冻土加速解冻并导致地貌失稳。这些物理变化直接改变了微生物赖以生存的基质条件——从水分可利用性、养分释放速率到氧化还原状态均发生显著波动,进而影响其群落结构、代谢路径与生态功能。
更为深远的影响在于生态级联效应与气候反馈机制。作为冰冻圈食物网的基础,微生物群落的变化可能通过自下而上的方式扰动整个生态系统,影响原生动物、微型后生动物乃至更高营养级的生物分布与丰度。同时,随着冻土融化释放出大量封存的有机碳,原本作为碳汇的冰冻圈可能转变为温室气体源。在厌氧条件下,产甲烷菌将有机质分解产生甲烷;而在好氧环境中,异养细菌则主要释放二氧化碳。这两种强效温室气体的逸出将进一步加剧全球变暖,形成正反馈循环。
Sugden, S., Davis, C.L., Quinn, M.W. et al. Current and projected effects of climate change in cryosphere microbial ecosystems. Nat Rev Microbiol (2025). https://doi.org/10.1038/s41579-025-01251-1
本综述系统梳理了不同冰冻圈生境的特征及其微生物群落的组成与功能,重点探讨了气候变化如何通过调控底物供给、酶动力学和微环境化学参数,在短期响应与长期演替两个维度上重塑微生物生态。此外,文章评估了此类生态转变对局部生态系统稳定性及全球气候系统的潜在影响,并指出现有研究中的关键知识缺口,旨在为未来预测模型的构建和应对策略的制定提供科学支撑。
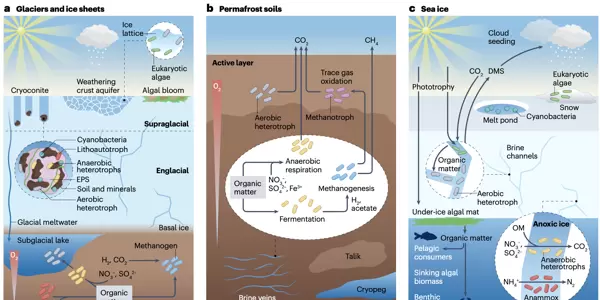
冰川和冰盖作为冰冻圈中最显著的组成部分,其微生物栖息地可依据空间分布划分为三个相互关联的子系统:冰上环境、冰内环境以及冰下环境。这些生态系统各自具备独特的物理与地球化学条件,并支持多样化的微生物生命活动。
冰上环境主要涵盖冰川表层的风化壳及表面形成的积水结构,如冰尘穴。风化壳是由冰面经历融化-再冻结过程形成的多孔结构,其下方可能发育含水层。而冰尘穴则是由于深色沉积物(俗称“冰尘”)吸收太阳辐射导致局部融冰所形成的孔洞,因其较高的生物活性常被视为冰川表面的生态“热点”。生活在此类极端环境中的微生物必须应对剧烈的日间与季节性温度波动、高强度紫外线照射以及水分供应的不稳定性。为适应这些胁迫,它们演化出多种策略,例如合成类胡萝卜素等光保护色素,或分泌胞外聚合物(EPS)以维持细胞稳定性和聚集矿物颗粒。
在营养极度匮乏的冰上系统中,初级生产者主要包括真核藻类(多见于风化壳)和蓝细菌(优势存在于冰尘穴)。它们通过光合作用固定二氧化碳,构成整个生态系统能量输入的基础。这些自养生物支撑着一个复杂的微型食物网,成员包括异养细菌、古菌、病毒、真菌以及微型真核生物(如纤毛虫和缓步动物)。值得注意的是,冰尘穴内部存在明显的微环境梯度:外部区域光照充足且氧气丰富,适宜光合作用和好氧呼吸;而核心部分因微生物呼吸消耗氧气,逐渐形成厌氧微区,支持发酵、反硝化及产甲烷等厌氧代谢途径。
近年来研究还发现,病毒在调控冰上微生物群落动态方面发挥关键作用。它们通过裂解宿主释放有机质,参与养分循环,并可能借助水平基因转移赋予宿主更强的抗逆能力。
冰内环境指冰川冰体内部的空间。由于生物量极低且采样困难,该区域是目前冰冻圈中认知最为有限的生态系统之一。一个长期存在的科学争议在于:检测到的微生物是否具有代谢活性,还是仅作为冰川形成时封存下来的休眠“历史档案”?尽管尚无定论,但多项研究表明,冰内微生物群落的组成不同于冰上和冰下系统,提示可能存在特定的选择压力或局部存活机制。若存在活性,这些微生物可能依赖矿物表面的薄水膜或高盐卤水通道,在低温条件下维持基本生命活动。
冰下环境涵盖基底冰、冰下湖泊和冰下沉积物,属于永久黑暗、低温高压的封闭生态系统。其化学特征受上覆冰川融水输入和下伏基岩地质组成的双重影响。出乎传统认知的是,这一环境并非生命荒漠。微生物群落可通过降解冰川运动过程中掩埋的古老有机质进行异养代谢。更重要的是,在缺乏光能的情况下,化能自养微生物成为主要的初级生产者——它们利用冰川研磨岩石产生的氢气(H),或氧化还原态的硫、铁化合物获取能量,并以此固定CO。此类过程不仅维系了冰下生态系统的运转,还可能导致甲烷等温室气体的生成与释放。
已观测到多个冰下湖泊中存在活跃的微生物群落,其水体与沉积物之间形成从有氧到无氧的连续梯度:湖水通常富含氧气,支持好氧呼吸;但当水流进入下伏沉积物后,微生物活动迅速耗尽溶解氧,促使反硝化、硫酸盐还原、产甲烷等厌氧过程占据主导。氮、铁、硫等元素在此过程中充当替代电子受体或供体,驱动复杂的生物地球化学循环。同时,冰下排水系统将微生物和溶解养分输送至冰川前缘,对下游生态系统提供重要支持。
在陆地环境中,与冰川毗邻的区域主要由多年冻土和冰川退缩后新暴露的地表(即冰川前缘)构成。其中,多年冻土土壤可分为两层:表层为季节性融化的活性层,以有氧代谢为主导,如有机质呼吸和微量气体氧化;下层为永久冻结的多年冻土层,其中封存着大量远古有机质。在此缺氧环境中,微生物依赖硝酸盐、硫酸盐或铁离子作为电子受体进行厌氧呼吸。当这些电子受体耗尽时,则转而启动发酵过程,并发生氢营养型和乙酸裂解型产甲烷作用。
多数多年冻土微生物栖息于微小的卤水脉络或较大的冷冻盐池中,这些液态水相因溶质浓度高而保持未冻结状态。这类常年不冻的局部区域被称为融区,为微生物提供了必要的生存介质。
海冰生态系统表现出显著的垂直分层与季节动态,无论是全年持续存在还是年际冻融循环的类型均如此。在早春至夏季,冰下和冰缘常出现藻类水华,推动初级生产力上升。随着光合作用受限和有机质(OM)积累,异养呼吸加剧并消耗氧气,导致厌氧条件发展,反硝化、硫酸盐还原和厌氧氨氧化等过程随之增强。
海冰内部的微生物活动主要集中于狭窄的卤水通道中。藻类在此进行光合固碳,为需氧异养菌提供有机碳源。许多海冰中的藻类和细菌还能产生二甲硫醚(DMS),该物质释放到大气后可促进硫酸盐气溶胶形成,进而增强云层反射率,影响气候反馈。此外,藻类衍生的有机碳沉降后可滋养下层的远洋生态系统,实现跨生境的能量传递。
冰川前缘是随着冰川退缩而新暴露出来的区域,其土壤发育尚不成熟,有机质含量较低,但含有大量由冰川研磨作用产生的矿物颗粒。在演替初期,这类土壤中的微生物群落以能够氧化铁、硫等无机物的化能自养菌为主,它们通过产酸等方式促进岩石风化,释放出可利用的营养元素。随着时间推移,光能自养微生物如蓝细菌和藻类逐渐定殖,不仅有助于固着松散的沙土,还为异养微生物提供了有机碳源。在缺乏维管植物的早期生态系统中,真菌在稳定新形成的有机碳方面发挥着关键作用。随着生态系统的逐步发展,微生物群落结构趋于复杂,较老的前缘土壤中可能出现甲烷氧化菌等类群,使其转变为甲烷的汇。
多年冻土指温度处于零度或以下、且持续两年以上的土壤或岩层。其上方覆盖着每年经历季节性冻融循环的土层,称为活性层。作为高度异质性的微生物栖息地,多年冻土的特性受到有机质含量、含冰量以及地貌特征(如冰楔多边形)等多种因素影响。微生物主要存在于未冻结的卤水脉或较大的咸水包体中,这些微环境被称为冷冻盐池。总体来看,微生物的多样性和丰度随深度增加呈下降趋势。活性层土壤中以好氧异养细菌和甲烷氧化菌为主要类群;而深层多年冻土则属于厌氧主导的生态系统,其微生物群落由生长缓慢、适应寡营养条件的类群构成,能利用硝酸盐、硫酸盐、三价铁等作为电子受体进行厌氧呼吸,或通过发酵途径获取能量。丰富的有机质——包括易降解与难降解成分——为上述代谢过程提供底物,其中发酵产物最终可被产甲烷菌转化为甲烷。此外,病毒及原生生物捕食者对多年冻土中微生物群落的更替和碳循环具有显著的“自上而下”调控效应。
海冰是由海水冻结形成的一种独特生境,内部遍布高盐度的卤水通道网络,为微生物提供了生存空间。海冰内的微生物群落表现出明显的垂直分层特征,并呈现强烈的季节性动态变化,与周围海水中的群落组成存在显著差异。真核藻类(尤其是硅藻)是海冰初级生产的主要贡献者,单位体积的生产力有时甚至超过开阔海域。这些藻类聚集形成的冰藻垫构成了极地海洋食物网的重要基础,其所固定的碳可通过海冰融化输入下层水体,支持整个水生生态系统的运转。部分海冰藻类还能合成二甲基磺基丙酸酯(DMSP),该物质可被细菌转化为二甲硫醚(DMS),后者参与云凝结核的形成,可能间接影响气候系统。藻华结束后,异养细菌(如γ-变形菌)迅速增殖,降解残留的藻类生物质。冬季光照减少,光合作用减弱,加之微生物呼吸耗氧,海冰内部可能形成局部厌氧微环境,从而推动反硝化、硫酸盐还原等厌氧过程的发生。
雪作为一种分布最广却最为短暂的冰冻圈生境,其微生物群落来源主要依赖大气沉降。新降积雪中的微生物通常来自远距离输送;而长期存在的雪盖(例如覆盖在冰川表面的积雪)则会发育出更具特色的本地群落,其中雪藻常占据优势地位,尤其在夏季更为明显。进入冬季后,随着藻类活性降低,依赖其衍生有机碳生存的真菌和异养细菌相对丰度上升。雪中的微生物不仅是当地生态过程的一部分,也是向冰川冰及其他冰冻圈环境传播微生物的重要种源库。
| 栖息地 | 平均温度 | 平均细胞密度 | 优势类群 | 主要代谢方式 | 氧气张力 | 常见研究地点 |
|---|---|---|---|---|---|---|
| 冰盖与冰川栖息地 | ||||||
| 冰尘穴 | 0.1 - 0.5 °C | 10? 细胞/克 | 假单胞菌门、蓝细菌门、拟杆菌门、酸杆菌门、放线菌门 | 光能自养、硝化作用、异养 | 好氧,存在缺氧微环境 | 斯瓦尔巴群岛、喜马拉雅山脉、南极半岛、麦克默多干谷 |
| 风化壳含水层 | ~0 °C | 10? - 10? 细胞/毫升 | 真核藻类、蓝细菌门、子囊菌门和担子菌门(真菌)、假单胞菌门 | 光能自养、异养、交叉喂养 | 好氧 | 马塔努斯卡冰川(美国阿拉斯加)、格陵兰冰盖 |
| 冰内冰 | < 0 °C | 10? - 10? 细胞/毫升 | 蓝细菌门、假单胞菌门、装甲菌门、厚壁菌门 | 无直接代谢活动证据 | NR | 斯托冰川(瑞典)、古里亚冰帽(青藏高原) |
| 基底冰 | < 0 °C | 10? 细胞/克 | 硫/铁循环菌、绿弯菌门、脱硫菌门 | 化能自养、产甲烷作用 | 微好氧 | 马塔努斯卡冰川(美国阿拉斯加)、泰勒冰川(南极) |
| 冰下湖 | < 0 °C | 10? - 10? 细胞/毫升或克 | 硫氧化菌、铁氧化菌、氮氧化菌、乙酸菌 | 化能自养 | 水体有氧,沉积物厌氧 | 默瑟湖和惠兰斯湖(南极)、东沙夫卡特拉湖(冰岛) |
| 陆地环境 | ||||||
| 活性层土壤 | 季节性融化 | 10? - 10? 细胞/克 | 放线菌门、酸杆菌门、假单胞菌门 | 好氧异养、微量气体氧化、甲烷氧化 | 好氧 | 斯托达伦沼泽(瑞典)、北坡(美国阿拉斯加)、青藏高原、阿克塞尔海伯格岛(加拿大) |
| 多年冻土 | -2 °C 至 -15 °C | 10? - 10? 细胞/克 | 假单胞菌门、放线菌门、发酵菌和产甲烷菌 | 硫酸盐呼吸、铁还原、硝酸盐呼吸、发酵、产甲烷 | 厌氧或微好氧 | 斯托达伦沼泽(瑞典)、北坡(美国阿拉斯加)、青藏高原、阿克塞尔海伯格岛(加拿大) |
| 冷冻盐池 | -6 °C 至 -11 °C | 10? - 10? 细胞/毫升 | 海杆菌属、嗜冷杆菌属、吉里西亚菌属、γ-变形菌纲、放线菌门 | 异养、硫酸盐还原、产乙酸、产甲烷 | 厌氧或微好氧 | 乌特基亚格维克(美国阿拉斯加)、亚马尔半岛(西伯利亚) |
| 冰川前缘土壤 | 季节性融化 | 10? - 10? 细胞/克 | 岩源自养型假单胞菌门、蓝细菌门、放线菌门 | 年轻土壤:化能自养和光能自养;年老土壤:异养 | 好氧 | 丹玛冰川(瑞士)、中洛芬布林冰川(斯瓦尔巴)、欧洲阿尔卑斯山脉、南极半岛 |
| 海洋环境与雪 | ||||||
| 环境类型 | 平均温度 | 平均细胞密度 | 优势类群 | 主要代谢方式 | 氧气张力 | 常见研究地点 |
|---|---|---|---|---|---|---|
| 海冰 | -2 °C 至 -20 °C | 10? 细胞/毫升 | 真核藻类(硅藻)、γ-变形菌门、拟杆菌门、放线菌门 | 光能自养、好氧及厌氧异养 | 好氧,冬季可变为厌氧 | 加拿大北极群岛、楚科奇海、罗斯海 |
| 积雪 | < 0 °C | 10? - 10? 细胞/毫升 | 真核藻类(绿藻)、γ-变形菌门、拟杆菌门、放线菌门 | 光能自养、利用藻类衍生底物的异养 | 好氧 | 冰岛、斯瓦尔巴群岛和南极的冰川表面 |
冰冻圈在全球碳氮循环中扮演着关键角色,储存了大量有机碳和营养元素。例如,多年冻土区域封存约1500 Pg的有机碳,而冰川与冰盖也固定了可观的碳与养分。随着气候变暖,这些储存库正面临多重扰动,微生物主导的生态过程随之发生显著改变。
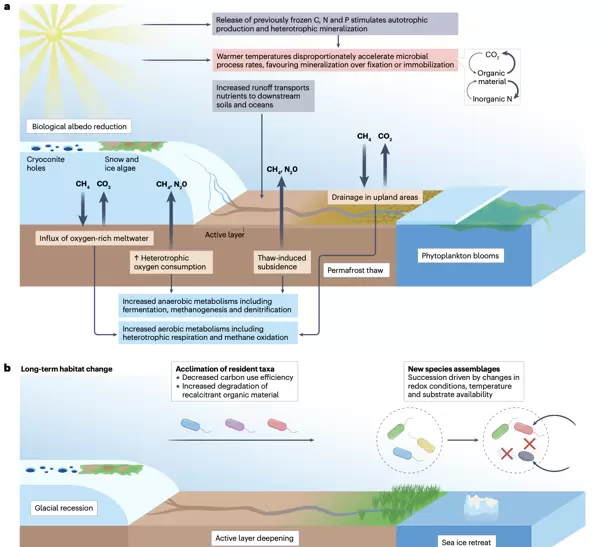
短期响应:底物释放、酶活提升与氧化还原波动
在较短时间尺度上,气温升高促使冰川、多年冻土及海冰加速融化,导致原本被冻结的碳、氮、磷等物质释放,从而提高环境中底物的可利用性(灰色路径)。同时,融水径流增强进一步向陆地和海洋系统输送养分与有机碳,扩大资源供给。
升温对生物化学反应速率具有非线性放大效应(粉色路径),尤其体现在酶活性的显著提升。矿化过程如异养呼吸和氨化作用的加快往往超过自养生产、固氮或氮固定等固定过程的速度。这使得多数冰冻圈生态系统中的初级生产与群落呼吸比值趋于下降,净碳释放增加。
此外,水文变化引发局部氧化还原状态转变(蓝色路径)。在冰下生境中,富氧融水流入可能暂时维持有氧条件;但若异养活动加剧且气体交换受限,则可能导致氧气耗尽,形成局部厌氧微环境。陆地上,冻土融化引起的地面沉降易造成土壤积水缺氧,而在排水良好的高地则可能促进通气,支持更多需氧代谢。
上述三方面——底物可用性上升、酶动力学加速以及氧化还原电位变动——共同重塑冰冻圈内各类栖息地的生态功能。例如,冰面藻华频率上升将降低反照率,加剧吸热;海洋浮游植物水华改变碳季节性分配;多年冻土解封释放大量有机质供异养降解。这些反馈机制推动CO、CH和NO等温室气体从历史汇转为大气源。
长期演变:栖息地丧失与群落更替
在十年至百年的时间尺度上,冰川退缩、海冰范围缩减以及多年冻土活性层加深,将导致大范围冰冻圈消失。原有微生物群落需适应更高温度和变化的资源结构,通常表现为碳利用效率降低——不稳定有机质优先消耗后,迫使微生物依赖更难降解的顽固性底物。
伴随物理环境的根本性转变,生态系统进入演替阶段,新的物种组合逐步建立。许多嗜冷类群逐渐被更能耐受融化后条件的种群取代,生态网络重构。这种长期动态不仅影响本地生物多样性,也深刻改变元素循环模式与温室气体通量。
底物可用性的提升
气候变暖直接引致冰雪与冻土解冻,释放出长期封存的有机质和营养盐(如氮、磷)。预计到本世纪中叶,多年冻土融化将释放巨量碳库,其中部分碳成分极易被微生物迅速利用。与此同时,冰川融水量上升增加了向淡水和海洋系统输入有机碳和养分的通量。
这些新释放的活性碳不仅能直接刺激异养微生物的代谢活性,还可能通过“启动效应”激活古老、稳定有机碳的分解过程。另一方面,养分限制的缓解也有助于光能自养生物(如冰面藻类、浮游植物)生长,提升初级生产力,进而为异养者提供更多可利用底物。
酶活性的温度敏感性增强
温度是调控生化反应速率的核心因素。即使在低温范围内,小幅升温也能显著提升酶促反应速度,加快有机质分解与养分转化。极地微生物过程通常表现出较高的温度敏感性(以Q10值衡量),意味着其对气候变暖响应更为剧烈。
值得注意的是,异养呼吸的Q10普遍高于光合固碳,这意味着随着温度上升,呼吸增长快于生产,导致生态系统净碳矿化增强,向大气释放更多CO。类似地,在低温条件下,产甲烷过程的Q10常等于或高于甲烷氧化过程,预示着变暖可能打破平衡,使CH生成超过消耗,形成净排放。
氮循环同样受到温度调控:矿化速率随温升而加快,缓解氮限制;但在低温下硝化作用缓慢,铵态氮积累。当温度继续升高,硝化与反硝化过程被激活,可能加剧氮素损失,并影响NO排放。
氧化还原状态的动态转换
气候变化不仅通过升温影响环境,还通过改变降水格局和融水流动路径,重塑栖息地的氧化还原状况,决定优势代谢途径是有氧还是厌氧,进而影响CH和NO的生成与消耗。
在陆域系统中,多年冻土融化诱发热喀斯特地貌,形成积水洼地,创造厌氧条件,利于产甲烷菌活跃。然而,若环境中存在铁氧化物或硫酸盐等替代电子受体,铁还原菌或硫酸盐还原菌会与产甲烷菌竞争底物,抑制甲烷生成。相反,在排水良好区域,融化改善通风,促进甲烷氧化菌活动。
在冰下生态系统中,持续增加的融水输入可能带来额外溶解氧,增强甲烷氧化潜力。同时,氧化还原条件的变化也调控氮循环:厌氧环境下反硝化作用加强,可能产生大量NO;而在好氧条件下,硝化作用占优,有助于氮素保留。
长期驯化与生态演替
面对持续变暖,微生物群落可通过生理适应(驯化)或种群替换实现长期生存。一些类群可能调整代谢策略以应对资源变化,而另一些则因无法适应而衰退。整体上,生态系统经历结构性重组,耐寒物种逐渐被适应温暖、湿润或富营养条件的新组合所替代。这一过程驱动冰冻圈生态功能的根本转变。
除了短期内对微生物生理活动的直接影响,气候变化还将引发微生物群落长期的结构与组成演变,这种变化带来的生态后果更为深远且难以准确预测。尽管在变暖初期可能观测到呼吸作用或产甲烷速率显著上升,但随着易降解有机底物的消耗,或是微生物群落逐渐适应新的温度环境(即发生驯化或功能适应),这些响应可能会减弱。有研究指出,升温诱发的异养微生物活性激增,可能导致后续出现碳源和养分的长期匮乏,迫使微生物转向分解更难降解的顽固性有机质,从而表现为碳利用效率下降,并伴随降解复杂碳化合物相关酶活性的增强。
最根本的转变源于生境本身的剧烈改变:冰川退缩、多年冻土解冻以及海冰消失,正在使原本占据主导地位的嗜冷微生物类群被更能适应温暖条件的物种所取代。这一过程不仅改变了群落结构,还可能彻底重塑生态系统功能——例如从碳汇转变为碳源。为了预测此类生态演替对未来地球系统的影响,科学家依赖时间序列监测与空间替代法进行研究,这些方法对于模拟和预估未来生态系统状态具有关键意义。
局部变化,全球影响

多年冻土融化
全球观测数据显示,多年冻土层温度持续升高,导致大量原本冻结的有机质解冻并暴露于微生物分解之下。野外调查与实验室培养实验均表明,在冻土融化后,微生物群落中与植物多糖降解相关的基因(如纤维素、半纤维素降解酶基因)丰度及表达水平明显上升。放射性碳定年结果显示,被矿化的碳中有相当比例来源于古老封存碳库,而非近期光合固定的现代碳,这直接证明了古老碳循环已被激活。与此同时,氮循环相关基因(如参与硝化和反硝化的基因)也发生变化,增加了N2O等强效温室气体的排放潜力。具体的代谢路径则受控于融化速度和局部水文状况:快速融化常形成热喀斯特湖,创造厌氧环境,促进产甲烷过程;而缓慢融化且排水良好的区域则更有利于好氧呼吸和甲烷氧化菌的活动。此外,冻土融化还可能释放历史上沉积的汞等污染物,这些重金属可在厌氧条件下经微生物转化为高毒性的甲基汞,并通过食物链富集,威胁生态系统和人类健康。
生物反照率降低
冰雪表面具有高反照率,能够反射大量太阳辐射,对维持地球能量平衡至关重要。然而,气候变暖延长了生长季节,改善了冰雪表面的水分、热量和营养供给,促进了富含色素的光合微生物(如雪藻、冰藻)的大规模繁殖。这些微生物合成的色素(如雪藻中的类胡萝卜素使其呈现红色或绿色)显著降低了地表反射能力,增强了太阳辐射吸收,进而加剧局部升温与冰雪融化,形成正反馈机制。
在格陵兰冰盖上,大规模冰藻水华已被证实可显著降低反照率,并增加融水径流。同时,冰尘穴的扩展进一步放大了这一效应。目前,生物性反照率下降已成为冰川质量损失模型中一个日益受到重视但尚未充分量化的重要因子。
浮游植物水华
在海洋冰冻圈区域,气候变暖引起海冰范围缩小、厚度减薄、表层海水温度上升,以及冰川融水带来的额外营养输入,共同推动极地海域浮游植物水华提前发生、持续时间延长、分布范围扩大。这种物候上的改变可能导致浮游植物水华高峰与浮游动物摄食高峰期“错配”,使得大量初级生产力未被有效利用而沉降至深海,影响能量向高营养级传递。
另一方面,海冰变薄提高了透光率,有利于冰下藻类的生长,这部分生产力在过去常被低估。更令人担忧的是,气温升高可能促使产毒藻类(如某些甲藻)扩散至新区域,并提高有害藻华的发生频率。其产生的毒素可在贝类等生物体内累积,并沿食物链传递,危及海洋生态安全和人类食品安全。此外,水华后期大量有机质的分解过程会消耗水中氧气,可能引发局部缺氧区,给海洋生物带来生存压力。
新兴挑战与展望
地理分布偏差
当前绝大多数关于冰冻圈微生物生态的研究集中于少数具备完善科研基础设施的地区,如斯瓦尔巴群岛、阿拉斯加和瑞士阿尔卑斯山区。然而,冰冻圈环境在空间上高度异质,不同区域(如格陵兰、西伯利亚、青藏高原)在变暖速率、地质背景和生态系统类型方面存在显著差异。因此,基于局部区域的研究成果难以外推至全球尺度。这种地理采样上的不均衡限制了我们对全球冰冻圈微生物响应模式的整体认知,也为模型预测引入了不确定性。
未来亟需加强对偏远、代表性不足地区的系统性考察,并积极联合当地社区(特别是原住民群体),整合传统知识与科学数据,构建更加全面、多样化的观测网络。
季节性偏差
目前多数冰冻圈微生物研究集中在夏季可进入期,导致对春秋季甚至冬季微生物动态的理解严重不足。事实上,冬季的生物地球化学过程(如低温下的缓慢分解、气体积累)可能对全年碳氮循环产生重要影响。同样,春季融雪期和秋季冻结初期也是关键过渡阶段,涉及微生物群落的快速重组与功能切换。因此,克服季节性观测盲区,实现全年度连续监测,是提升预测精度的关键方向。
在全球变暖的背景下,冰冻圈作为独特的微生物生态系统,正面临前所未有的变革。气候变化不仅通过直接途径——如释放冻结养分、加快代谢速率、改变生境的氧化还原状态——影响微生物活动,还通过长期驱动群落结构的根本性演替,深刻重塑其生态功能。这些变化不仅威胁冰冻圈内部生态稳定与生物多样性的维持,更通过温室气体排放、海平面上升、地表反照率改变、水资源重新分布以及对食物安全的影响,引发区域乃至全球尺度的连锁反应。
尽管如此,当前相关研究仍存在显著局限。地理覆盖不均、季节性数据缺失,尤其是缺乏长期动态观测记录,严重制约了我们对微生物响应机制的理解。特别是冬季,作为气候变暖效应最明显的季节,其微生物活动的研究尤为薄弱。由于极端环境与后勤困难,大多数野外工作集中在夏季,导致我们对漫长极夜期间微生物的行为几乎一无所知。然而,越来越多的研究表明,即使在亚零度条件下,微生物仍可能保持代谢活性;积雪或海冰形成的隔热层甚至能创造出高于冰点的微环境。这意味着冬季微生物过程对全年碳氮循环通量的实际贡献可能被严重低估。

为了弥补这一知识空白,亟需发展自动化的原位监测传感器,结合遥感技术,并推进可在实验室模拟冬季条件的培养实验。此外,仅依靠横断面调查或短期观测无法揭示微生物对持续升温的非线性响应及其长期演替规律。因此,建立长期观测站点,实施原位增温措施(例如使用开顶箱或雪栏调控),并辅以受控环境下的室内模拟实验,成为关键手段。这类长期操控实验有助于识别微生物群落的适应能力,并发现重要的生态阈值——比如系统从碳汇转变为碳源的关键节点。
唯有整合长期观测数据、实验结果与数学模型,才能显著提升我们预测冰冻圈生态系统未来演变的能力。面对快速变化的气候格局,必须推动跨学科协作,融合前沿技术与多尺度模拟方法,深入解析微生物在冻土、冰川、海冰等环境中的作用机制。这不仅关乎极地生态保护,更是应对全球气候变化、保障地球系统可持续运行的重要科学支撑。
关注公众号 ‘bio llbug’,获取最新推送。

 京公网安备 11010802022788号
京公网安备 11010802022788号







